Morphology and Anatomy of Bryophyta - Mosses
Last updated: 16.10.2021
The mosses, systematically referred to as Bryophyta, are with around 13 000 species not only the largest division of the bryophytes, but also the second largest division of the land plants [1, p. 55]. Except for the sea, they have conquered all habitats on earth, from tropical rain forests to the poles, from deserts to alpine regions. According to molecular phylogeny, mosses split from other plants in the age of the Ordovicium, more than 440 million years ago. Yet, unambiguous fossil records exist only from the upper carboniferous onwards, more than 300 million years ago [3] [1, p. 55]. In more than 400 million years of evolution, several evolutionary radiations have occurred, which established a wealth of morphological, developmental, anatomical and cytological diversity. Actually, this wealth is so vast no fully applicable description of the mosses can be given, exceptions to almost every characteristic are found. Difficulty is increased further by reductions that have occurred. Most mosses show a seta carrying the capsule, which has one or two rows of teeth (the peristome) at the opening (annulus). This sporophyte remains connected to the gametophyte for his entire life and is at least partially nourished by it. The gametophyte often consists of a round stem with sessile leaves and is always self-autotrophic [1, p. 55] [2, p. 70]. The connection between Bryophyta and the other divisions, Marchantiophyta and Anthocerotophyta, is their heterophasic and heteromorphic life cycle [4, p. 2].
This chapter is intended to provide an overview of the development, morphology and anatomy of the mosses, as well as some information about reproduction and life cycle. It starts with the general developmental plan of mosses, followed by a more detailed description of the important structures: in the gametophyte rhizoids, stems, leaflets, and modifications of these, such as branches, their arrangement, as well as the sexual organs; in the sporophyte development, placenta, capsule, spores and their formation, the peristome.
Notes on progress: The section on general characteristics of the gametophyte is finished, but will be updated with more detailed information soonTM. Espescially pictures and diagrams are missing. The sporophyte is in progress. Systematics were outsourced to its own section.
The modular growth of mosses
The structure of the gametophyte, the actual moss plant, can be regarded as modular. A single apical meristematic cell forms cells that develop further into metameres and modules . Usually, the apical cell is three-edged and corresponds to an inverted tetrahedron. On all three sides of the tetrahedron cells are separated in a clockwise fashion, which are then divided through a pericline cell wall into an inner and outer daughter cell. The inner cell goes on to produce the internal tissue of the stem. The outer cell, which protrudes from the surface in a papillary fashion, again divides periclinary. The outermost cell will go on to form the leaves, the inner one the epidermis. A metamer has formed. All metamers of an apical cell together form a module. Growth in the longitudinal direction is achieved by cell division, cell enlargement and cell extension. Vegetative growth is the result of a build-up of these cells, all of which are derived from the apical cell. [1, p. 56] [2, p. 70 et sqq.] [5, p. 83] [8, p. 12]
Branches can occur when a three-edged initial cell is formed from a cell of the epidermal tissue . Usually, the so called branch initial divides to form a primordium, which goes on to develop into a branch. If the primordium falls dormant after developing tiny leaves a bud has arisen. Without leaves the structure is called a naked primordium. Since this development starts with the cell below the initial cell for leaflets, a branch is always below the leaflet of the corresponding metamer. The branch consists of exactly the same structure of metameres, modules and intital cells for leaves and branches. Thus, branches can themselves branch. Not all initial cells seem to be active, because not all mosses form branches. However, foliage is common. [1, p 56-60] [2, p. 71 et sqq.] [5, p. 83]
There are also deviations from this basic blueprint. The genus Fissidens has double-edged apical cells in the form of a biconvex lens. But these, too, are based on a tetrahedral cell in their development. In Sphagnum, the apical cell is supported by a meristem located below the cell. In Takakia lepidozioides, meristematic activity is taken over by a ring of cells. In the middle of it lies a silent central cell [1, p. 56].
Branching and Gametangia
As can be seen from the above description, any metamer can produce a branch. The density of branches, determined by dormancy of the initial cell, is not the only variation that can occur.
- Location: Basitone/Basipedal branching takes place at the end of the module, new branches are thus below older ones. Acrotone/Acropetal branches start at the top of the module, new branches are above old ones.
- Mode: If a new stem emerges from a branch that is identical in function to the old one, its called sympodial branching. The system of metamers is replicated. This occurs, for example, in species where the development of a gametangium makes the extension of the original stem impossible. With monopodial branching, the formation of the sex organs is transferred to branches. This means, the stem can basically grow infinitely. Branching of branches can produce tertiary and even quaternary branches.
- Direction: An orthotropic branch grows orthogonal, perpendicular, to the substrate. A plagiotropic branch is parallel to the substrate.
Type of branching and location of the gametangia is key for an important clasifiacation of the mosses, namely into acrocarpous and pleurocarpous mosses. Acrocarpous mosses form the gametangia at the top of the stem, whereby the apical cell is consumed. Pleurocarpous mosses show monopodial branching. The stem can grow almost infinetly, since the formation of gametangia has been transferred to specialized branches. A variation of pleurocarpy are the cladocarpic mosses, in which each branch is fully developed. Most of them end with female gametangia. A systematic relevance of this branching pattern is not assumed [6, p. 174]. Pleurocarpous mosses seem to have emerged from acrocarpous ones on several occasions. [1, p. 72 et sqq.] [2, p. 72]
The Gametophyte
The gametophyte represents the actual moss plant. Macroscopically it can be divided into rhizoids, stems (cauloids) and branches and leaflets (phylloids).
Rhizoids
Rhizoids can be understood as the bryophytes analogue to roots. Like roots, they anchor the gametophytes to the substrate and are involved in water conduction [1, p. 57].
Development and morphology
The growth of rhizoids begins with epidermal cells in the lower part of the stem, on the stems ventral side and/or also on the costa. Rarely are they formed at the apical part of the lamina by specialized nematogens (initial cell of rhizoids). In many mosses rhizoids are seldom observed in nature, often however under laboratory (in vitro) conditions. It can be inferred that formation is determined by environmental conditions, but the exact factors are not known. Likewise, density, length and branching is dependent on the substrate. Takakia never has rhizoids. In Sphagnum they occur only on the protonema and never on the adult gametophyte, except for one species native to New Caledonia. [1, p. 57]
The structure of rhizoids can be described as multicellular yet often uniseriate filaments of elongated cells with oblique cell walls. The surface can be smooth or papillose, the color can range from white to brown and violet. [1, p. 57] [2, p. 81] [5, p. 93]
Functions
The anchoring effect of rhizoids is due to their thigmotropic growth, meaning they wind tightly around objects in the ground. This behavior is particularly pronounced in some Polytrichaceae, in which small rhizoids grow around larger ones forming a spiral. Exceptions to this growth pattern are the rhizoids of Sphagnum, Andreaea and Andreaeobyrum. In some mosses that grow on soil, the rhizoids form a dense white to reddish brown felt around the stem . Its purpose could be that it conducts and/or stores water through capillary forces. In some Mniaceae, strongly branched rhizoids are formed around the initial cell of a branch. A protective function is assumed.[1, p. 57] [2, p. 81].
Gemmae and tubers
In some species rhizoids develop brood bodies, which are called gemmae or tubers. Gemmae are a kind of vegetative brood body consisting of a few cells that look similar to buds, leave or branches. However, they can also be formed by thalli, stems, leaflets. Under favorable conditions, they differentiate into a gametophyte [7, p. 96]. Tubers are similar to gemmae, but are only formed by rhizoids, usually underground [7, p. 208]. Another difference is the absence of abscission cells as they occur in gemmae. Tubers can be single- to multicellular, roundish or consist of a row of cells. Their development can be triggered by drought. Whether they actually survive longer periods of drought is not proven. They are developed by various acrocarpous Mosses such as the Polytrichaceae, Funariaceae, Bryaceae, Pottiaceae and Dicranaceae, but are apparently missing from the pleurocarpous ones. An infection of rhizoid cells with fungi can lead to formation of appendages that look similar to tubers [1, p. 57 et sqq].
 |
_8818.jpg) |
Photo by HermannSchachner, CC0 |
Photo by HermannSchachner, CC0 |
Stems
The term cauloid, invented to destinct the structure from the stems of cormophytes, is used less frequently [4, p. 77].
Basic anatomy
The cross section can be round, oval, or have three or more (weakly pronounced) edges. These edges often arise from adhesion of leaves. from the ground up the diameter first increases somewhat, then stays the same. Diameters range from 0,1 mm to 1,4 mm. From the outermost to the innermost layer, a stem often consists of epidermis, sterome or outer cortex, inner cortex and central strand or hadrom. Different degrees of differentiation can be observed . The terms used for the different tissues in stems are not uniform . [1, p. 58, 61] [4, p. 77] [5, pp. 83-84] [8, p10].
-
Epidermis: The epidermis can be comprised by a single to multiple layers, the transition to the cortex gradual or abrupt. Stomata are never present, but a cuticula, if only thin, can be observed. One variant is the hyalodermis, in which the cells are significantly enlarged and have thin cell walls, sometimes with spiraling reinforcement. In Sphagnum, sometimes pores are found in the wall .
-
Stereom: Also called external cortex, a single- to multi-cellular layer with strongly thickened cell walls. The cells are elongated (prosenchymatic), with very narrow lumen. Sometimes, these are called stereids. These stereides retain their protoplast even if mature and are colored brown, red or yellowish through pigmentation. The cell walls contain a polyphenolic compound, which is not lignin. The hollow cylinder formed by the stereome supports the plant.
-
Inner cortex: Formed by parenchymatic tissue with elongated cells whose ends are flat or roundish. The cells can store lipids or starch and are involved in the transport of water and nutrients through the plant.
-
Central strand: Also called Hadrom. Consists of elongated cells with a narrow lumen. Especially pronounced in acrocarpous species.
The degree of differentiation of the stem varies from species to species. Specialized strands for conduction of water and nutrients can be observed, which are called hydroids and leptoids. Generally, in Polytrichopsida the highest degree of differentiation, in the Andreaopsida the lowest degree of differentiation is found [1, p. 60] [4, p. 77].
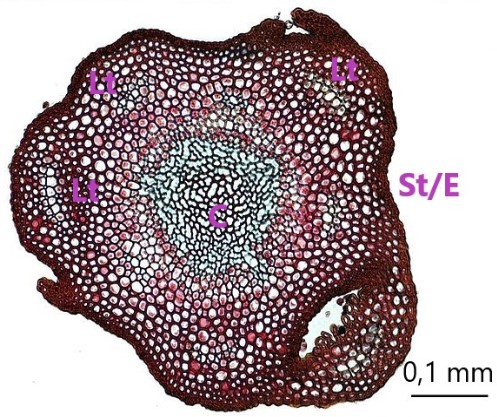
Stem cross sections showing various degrees of differentiation, interpretation with [5, p. 83 et sqq.]
 |
_7667.png) |
| Polytrichum spp. C marks the central hadrom, sorrounded by leptoids. Lt are some of the visible leaf traces, St/E the sterome and epidermis Photo by Jon Houseman / CC BY-SA, modified. |
Catoscopium nigritum. C is the central hadrom, sorrounded by a cortex Co without further differentiation. Lt marks a possible leaf strand which terminates in the epidermis E. Photo by HermannSchachner / CC0, modified. |
_7984.jpg) |
_1071.jpg) |
| Syntrichia ruralis. Basically no differentiation beyond a central hadrome C and an epidermis E. Photo by HermannSchachner / CC0, modified. |
Sphagnum capillifolium. The epidermis E is surrounded by a thick hyalodermis H composed of large cells with thin cell walls. No differentiation of the central strand C. Photo by HermannSchachner / CC0, modified. |
Transport of water and nutrients
Within the gametophyte transport of water and nutrients is necessary. For example, developing metamers must obtain assimilates and probably other organic compounds to cover their energy expenditure from other metamers or even modules. These transport tasks are carried out by undifferentiated parenchyma cells or specialized conductive tissues [1, p. 61 et sqq.].
Hydroids
Hydroids are cells specialized in the conduction of water. They are similar to stereids due to their prosenchymatic shape, but they have no protoplastic content. Also, the thickness of the cell wall is variable, it can be thin, or laterally reinforced with thin ends. Unlike trachids of vascular plants, no lignin (but other polyphenols), no annular or spiral cell wall reinforcements and no xylans are observed. The cell wall chemistry is very diverse. Together with associated parenchyma cells and stereids the hydroids form the totality of water-conducting tissues, the so-called hadrom. The hydroids of Takakia are built differently. They are short with right-angled ends and a differing cell wall composition. However, plasmodesmata are found, which are missing in Polytrichopsia and Bryopsida. It is therefore assumed that hydroids are not homologous structures [1, p. 61]. In Sphagnopsida, Andreaeopsida and Andreaeobryopsida no hydroids are observable.
Leptoids and conductive parenchyma cells
Nutrient-transporting cells are elongated, possess end walls with numerous pores derived from plasmodesmata, they lack vacuoles and the nucleus is degenerated. These similarities with sieve cells of the vascular plants indicate specialization for nutrient transport. However, experiments indicate a low level of optimization. This differentiation is most strongly pronounced in the Polytrichales and only in these the term leptoids is used. In all other taxons they are referred to as conductive parenchyma cells. Overall, nutrient-transporting tissues are difficult to identify in light microscopy, since many of their features are found in many other cells. In the stem one can find them throughout the cortex, sometimes with a preference for regions around the hydroids [1, p. 62].
Leaf traces
Sometimes leaf traces composed from hydroids and nutrient-conducting cells are observed. Their thickness and connection to the central strand varies greatly between and also within a species. There is a dependence on the water availability in the environment. Genuine leaf traces are mainly found in the Polytrichaceae, while false traces are characteristic of the Bryopsida [1, p. 62]. Interestingly, Frahm (2001) [4] denies the functionality of leaf traces in any species, while other sources like Goffinet (2009) claim they do work in Polytrichum.
Central cavity
In two tree-shaped hypopterygiaceae, namely Canalohypopterygium commutatum and Catharomnium ciliatum, cavities are found in the stem. It is believed that this cavity is involved in water transport or water storage . Whether it arises from schizogeny or lysogeny is not entirely clear. Lipids are found in the cavities, whose purpose could lie in energy storage or as a defense against herbivores or infections. [1, p. 63] [4, p. 77].
Paraphyllia, hairs and pseudoparaphyllia
The stem of a moss can be covered with various outgrowths different from leaves, branches, or rhizoids.

Paraphyllia of Thuidium delicatulum.
Patrick Alexander, CC BY-NC-ND 3.0
Paraphyllia are found on the stems and branches of many pleurocarpous mosses. They are small epidermal outgrowths which form ranges from single to branched filaments to leaf-like. They differ from rhizoids in their smaller cell size and green pigmentation. Their function lies in external capillary water conduction (when formed in large numbers like in Thuidium) and photosynthesis [1, p. 58, 4, p. 78]. At the insertion of a leaf into the stem, axillary hairs formed by the leaves are found. In pleurocarpous mosses they can also cover the whole stem. They usually consist of a unicellular unbranched filament of hyaline cells, seldom only one cell. The apical cell tends to be longer and shaped like a club, while at the base one or more brown cells can be found. Size, number, and shape show wide variations between species, they could thus be interesting for taxonomy and phylogeny. Their usage is however limited. Axillary hairs located at the leaf insertion secret mucilage. Their function could thus lie in preventing desiccation of young apices. The function of the hairs scattered around the stem is unclear [1, p. 58 et sqq.].
In genera like Dicranum and Encalypta another type of hairlike structures can be found on branches. However, these trichomes are seen as a type of mucilaginous leaf since they arise from a different metamer than the leave below them, unlike axillar hairs. In Andreaeobryum and Takakia beaked mucilage or slime papillae are found [1, p. 59].
The term pseudoparaphyllia is used for different structures by different authors. Traditionally it describes filamentous to foliose appendages in the immediate vicinity of buds and branches. Their function is the protection of the primordia. Akiyama & Nishimura (1993) distinct pseudoparaphyllia which arise from the stem and scaly leaves formed by the bud itself. Ignatov and Hedenäs (2007) use the term for all appendages near leaf insertions, primordia and the ones scattered along the stem. It also includes structures which were usually called paraphyllia. They use paraphyllia only for structures arranged in longitudinal fashion. The use in phylogeny is hampered by controversies about their homology [1, p. 60].
Pseudopodia

Pseudopodium with undehisced capsule of Sphagnum sp.
Wlodzimirez, Public domain, via Wikimedia commons
If a module forms female gametangia called archegonia at its apex, the apical cell is consumed and growth of the gametophyte ceases, except in Sphagnopsia, Andreaeosida and some species of Neckeropsis. In these growth starts again after fertilization of the archegonium, the so called pseudopodium elevates the capsule [1, p. 63].
In Andrea growth is the result of meristematic activity of the archegonial stalk. Cauline tissue seems to be involved as well, since reduced leaves, axillary hairs and archegonia can be observed on the pseudopodium in some taxa In Sphagnopsida the pseudopodium is formed solely by the cauline tissue. There is speculation both taxons acquired their pseudopodium independently of each other [1, p. 63].
The pseudopodium of Aulacomnium is a leaveless extension of the stem which carries at its apex brood bodies arranged in a crown. If this extension is formed by apical or subapical meristem is unknown [1, p. 63].
Leaves
All species of mosses form leaves, which distincts this group from hornworts and liverworts. Mosses developed a wealth of different leaf morphologies, lamina cells, costas, margins and more. The sole unifying character of all moss leaves is the absence of a stem. All leaves are sessile and inserted into the stem among their entire base [1, p. 64]. The many different morphologies also explain why mosses are mostly diagnosed by their leaves [4, p. 79]. Leaf morphology shows adaptions to the poikilohydric life of mosses. Cuticles are missing or reduced to not impede hydration. The lamina is undifferentiated and often unistratose, which improves gas exchange with the atmosphere.
Here, development and special orientation of leaves on the gametophyte will be covered, followed by morphology of the leaves: lamina, costa, margin, and ornamentation.
Development
During the development of a metamer started by the apical initial cell a single leaf initial cell per metamer arises. It is usually two-sided and produces derivatives on both sides in an alternating fashion, but only in one plane. The initial only produces six derivatives in each direction before it ceases to divide. Thus, the lamina arises from divisions of these derivatives. The pattern and timing of these divisions is different between the parenchymatous networks of five to six sided cells found in Polytrichopsida and most acrocarps and prosenchymatous networks of elongated cells of most pleurocarps. The wide rhombic cells of the Bryaceae and Hookeriaceae is assumed to have a transitory position between parenchymatous and prosynchomatous networks [1, p. 66 et sqq.]. In parenchymatic networks, dividing cells arrange the spindles separating the chromosomes in a 90° angle. This results in isodiametric cells. Prosenchymatic networks start out with rhomboid cells which through expansion of their cell walls morph into elongated cells with pointy ends [4, p. 82]. Multistratose laminas and costas are the result of a series of periclinal and anticlinal divisions, which leads to several median cell layers [1, p. 67].
In Andrea the leaf initial can have one or two cutting faces. In Buxbaumia, with its reduced leaves, no apical cell is found. If instead a basal meristem is responsible for leaf formation is unknown [1, p. 67].
Spatial orientation and foliar dimorphism

Leaves forming a dense rosette in
Rhodobryum roseum
Kurt Stüber, CC BY-SA 3.0, via Wikimedia Commons
The angle between consecutive leaves is the result of the geometry of the apical cell. A perfect tetrahedron would produce metamers in three directions, 120° angled towards each other. Since a full rotation has 360°, every fourth leaf would be aligned vertically. However, such phyllotaxis is not observed in gametophytes. Close examination of the apical cell reveals that the angle between metamers is about 137°. Only every seventh leaf would align. In nature, mostly obscure spiralling is observed. This maximizes light exposure and minimizes shades [1, p. 64 et sqq.]. In some taxa, e.g., Conostonum, the leaves form five ranks. Fissidens with their lenticular apical cell, but also in Bryoxiphium and Distichum with normal tetrahedral apical cells, arrange their leaves in distichous fashion. Shistostega starts with radially inserted leaves at the apex, but torsion of the stem leads to a distichous arrangement further down the stem [1, p. 65]. In plagiotropic mosses living in low light conditions coplanate arrangement is observed. The leaves are inserted in more than two ranks but in maturity all of them are alinged in a single plane. One example is Plagiothecium [1, p. 65].
The distance between leaves depends on the elongation of the metamer. Most mosses show a more or less regular foliation. In non-photosynthetically active stems the spacing is wide. In Rhodobryum metamers near the apex elongate only sparingly, leading to a dense rosette of leaves at the apex and wide spacing below [1, p. 66].
 |
_1777 CC0 HermannSchachner.JPG) |
 |
| Spiral arrangement in Polytrichnum commune James Lindsey, CC BY-SA 2.5, via Wikimedia Commons |
Distichous arrangement in Fissidens adianthoides HermannSchachner, CC0, via Wikimedia Commons |
Coplanate arrangement in Plagiothecium denticulatum Kristian Peters, CC BY-SA 3.0, via Wikimedia Commons |
Foliar dimorphism
In several instances different leaf morphologies correlating with the position of the leaf on the gametophyte are observed: Heteroblastic series, anisophylly, heterophylly, functional dimorphism and more.
In a heteroblastic series leaf morphology changes along the stem. A juvenile leaf at the basis of a module shows a different morphology compared to a juvenile leaf at the apex and any of its developmental stages. Heteroblastic series are common in mosses, but the difference is often minute. In distantly related mosses similar juvenile leaf morphologies can be observed, even though mature leaves are very different, e.g. Tortula and Funaria. This illustrates the phylogenetic usefulness of this phenomenon [1, p. 65].
Anisophylly describes differing leaf morphologies between different modules of a plant. In Macromitium leaves on creeping stems are reduced compared to the ones on erect branches. Some mosses are organized similar to a fern, with a vertical stipe and horizontal rachis. In these, the leaves on the stipe are scalelike and without a midrib, while the ones on the rachis are fully developed with a midrib [1, p. 65].
Heterophylly refers to leaf morphology dependent on in its position on the circumference of the stem. Reduced ventral or dorsal leaves compared to lateral ones is rare in mosses and occur e.g., in the Hyperpterygiaceae. In plagiotropic mosses like Racopilum this increases light exposure of the lateral leaves. Why orthotropic mosses like Epipterygium developed this adaptation is unclear. Amphigastria, reduced ventral leaves, can fulfil other functions besides photosynthesis like water retention. In Caytophorum tahitense the bases of amphigastria forms a little pouch to hold water.
Functional dimorphism arises when certain leaves are adapted for special purposes. In Calymperaceae stenophyls carry gemmae for asexual reproduction. Their lamina is reduced, the costa rod-like. Splash-cups of dense rosettes are formed around antheridia in Polytrichaceae and some Mniaceaea and discoid gemmae in Tetraphis. Differing leaf morphology is also often observed around the gametangia. Around the antheridia of autoiocous mosses leaves are similar to juvenile ones and thus much smaller than the others. The leaves around archegonia (perichaetial leaves) are often larger and longer than the rest. Most often, the innermost perichaetial leaves are the largest (e.g. in Hypnales), sometimes they are the smallest (e.g. in Mniaceae) [1, p. 66].
Lamina and lamina cells
The leaf is where mosses show most of their morphological diversity. An exhaustive account is not possible. In general, the lamina is unistratose, with exception of the costa and sometimes marginal regions or longitudinal strands (e.g. in Grimmia or Orthotrichum). Stomata are never found, cuticles can be present, but usually are so thin that they can’t effectively stop water loss [1, p. 67].
Outline and margin
The leaf is usually a single, continuous blade with an orbicular, to lingulate truncate to linear outline.
Overview over diffrent moss leaf outlines, for all images HermannSchachner, CC0, via Wikimedia Commons
_2525_barb.jpg "Anoectangium_aestivum_%28b,_113311-471220%29_2525_barb") |
_1493_bearb.jpg "Atrichum_tenellum_%28b,_150453-482253%29_1493_bearb") |
_4446_bearb.jpg "Cyrtomnium_hymenophylloides_%28b,_145601-473026%29_4446_bearb") |
| Anoectangium aestivum | Atrichum tenellum | Cyrtomnium hymenophylloides |
_2650_bearb.jpg "Dicranella_howei_%28a,_153802-482539%29_2650_bearb") |
_6958_bearb.jpg "Physcomitrella_patens_%28a,_144649-481016%29_6958_bearb") |
|
| Dicranella howei | Physcomitrella patens |
Exceptions are Takakia, Buxbaumia, Schistostega and Fissidens: Takakia has deeply segmented leaves, as are the perigonial leaves of Buxbaumia. In fertile stems of Schistostega leaves can be connected by their lamina at the base. Despite the appearance of a continuous lobed ribbon it is formed by individual consecutive leaves [1, p. 67]. The leaves of Fissidens have a Y-shaped cross section, at least at their base. The arms are the vaginant lamina, the base the dorsal lamina. All three connect to the costa in the middle. The vaginant lamina spans half to ¾ of the leaf’s length, then a single ventral laminal blade as well as the dorsal lamina continues the leaf. The dorsal lamina runs the entire length of the leaf but is often not inserted into the stem. In rare cases it is decurrent. The vaginant laminae can be seen as the true leaf, while the ventral and dorsal laminae are just outgrowths of the costa [1, p. 67].
The margin of the lamina can be entire, dentated, serrulated, serrated or laciniaeted/ciliated. Dentations consist of conical projections of the cell wall, serrulations of cell apices. Serrations consist of a single whole cell while laciniae or cilia are comprised of multiple cells in a hairlike structure. (Differing, [7, p. 178] uses serration for protrusions of one or more cells. Dentation refers to outward facing teeth consisting of one or more cells [7, p. 63]). Besides laciniae all are common.Teeth can be single or in pairs, span the entire margin or only a part of it [1, p. 67]. In Mnium, the margin is formed by a band of elongated cells, two or more cells wide. They could be important for water conduction or for protecting the structural integrity [1, p. 69]. Sometimes the margin is multistratose (Mniaceae) [1, p. 67] or curled in towards the midline, either in an upwards or downwards direction.
All pictures by Hermann Schachner, CC0, via WikimediaCommons. Editied by Simon Hofer
 |
_2940.JPG "Funaria_hygrometrica_%28e,_144437-481001%29_2940") |
_4450.JPG "Timmia_norvegica_%28a,_133246-471443%29_4450") |
| Smooth entire leaf margin of Bucklandiella microcarpa | Serrulations on Funaria hygrometrica | Small tooth (serration) on Timmia norvegica |
_8594.JPG "Timmia_austriaca_%28a,_145612-473017%29_8594") |
_9424.JPG "Mnium_spinosum_%28a,_113302-471423%29_9424") |
_8668.JPG "Bryoerythrophyllum_recurvirostrum_%28b,_144741-480322%29_8668") |
| Strongly pronounced serration on Timmia austriaca | Strongly pronounced margin of elongated cells on Mnium spinosum | Downwards currled margin of Bryoerythrophyllum recurvirostrum |
Lamina cell shape
As mentioned in the section about development, lamina cells are grouped according to their shape into parenchymatic, prosenchymatic and intermediate lamina cells. Parenchymatic cells are isodiametric, for example square, rectangular, round. They are found in Polytrichidae, Tetraphidiae, Buxbaumiidae, Dicrananae, Orthotrichanae, Bartramianae. Prosenchymatic cells are often much longer than thick, with ratios up to 10 to 1. The shape can be rhomboid, long hexagons or irregular with drawn out peaks [4, p. 82 et seqq.]. Among length and width of a leaf lamina cells can change their shape. Sometimes a differentiation into small isodiametric, chlorophyllous upper and bigger rectangular and hyaline cells is seen, e.g., in Encalyptaceae, Pottiaceae, Calymperaciae. In most pleurocarps alar cells are found, which are small, dense and quadratic or large, thin walled and hyaline (diagnostic for Sematophyllaceae). Their purpose could be absorption of excess water to slow desiccation. If mosses are grown submerged, their differentiation is reduced. However, another effect of water absorption is an increase in turgor, which can push the leaf away from the stem [1, p. 69, 4, p. 84 et seqq].
Ornamentation

Examples of papillae, Orthotrichum lyellii (above)
and Encalypta streptocarpa (below).
Pictures by Hermann Schachner, CC0, WikimediaCommons.
A very striking property of mosses is the ornamentation of lamina cells seen in some taxa. This ornamentation can be grouped in mammillae, which are protrusions of the cell lumina, and papillae, which consist of cell wall. Papilla show a variety of structures: small knobby to large antler-like, cylindrical to crescents. Numbers range from one to several per cell. Presence of papillae (or a cuticle) gives the leaf a matte gloss [1, p. 69]. Their purpose is not entirely clear. Frahm (2001) claims they help break up incoming raindrops, thus decreasing time to turgescence. In Goffinet (2009) the increase of surface area is mentioned to, but another aspect is highlighted. A water film on a leaf significantly slows down diffusion of CO2 into the leaf. If the tops of papillae are covered by water repellent cuticle they can function like a snorkel; penetrating the water film and easing diffusion of gasses [1, p. 69, 6, p. 85].
Cell walls and leaf orientation
Cell walls can also be differentiated. Racomitrium has irregular wavy cell walls. In Dicranum the thick axial walls are porous, in Steyermarkiella the transverse walls of the hyalocysts [1, p. 69].
Differences in cell wall thicknesses also accounts for the leaves three-dimensional shape as well as movement when wetted. If the dorsal and ventral cell walls have different length and thickness along the whole leaf, a concave or convex shape results. Longitudinal folds called pleads or plicae and transversal folds or waves called undulations are due to locally confined cell wall differences [1, p. 70]. Strongly or randomly instead of crosswise undulated leaves are called rugose leaves [7, p. 174]. Why mosses acquired these folds is not known, but in some species they are lost when cultivated fully submerged [1, p. 70].
In Dawsonia vertical lamellae control the bending of the leaf when wetted: The thick cell walls hydrate and exert pressure. The transversely elongated and swollen cells are not involved in guiding the movement, rather they mechanically reenforce the leaf to prevent tearing. Hygroscopic movement is seen in many species. In Helicophyllum drying leaves curl in staring from the apex downwards [1, p. 70].
Costa
The costa or midrib is a multistratose structure running along the midline of a leaf. Usually, the costa is single and unbranched. However, it can be double, like in many pleurocarps, or missing entirely [1, p. 68]. In some fossils, three midribs can be seen [4, p. 80]. In very few taxa like Antitrichia it appears to be truly branched. Length varies from barely visible to reaching beneath the lamina, forming a toothed or smooth awn or hair [1, p. 68]. The purpose of a hyaline hair is ambiguous. It could facilitate condensation of moisture, or reflect sunlight to reduce evaporation [4, p. 80].
Pictures by Hermann Schachner, CC0, via WikimediaCommons
_3076.JPG "Hedwigia_ciliata_%28f,_153458-482459%29_3076") |
_9422.JPG "Syntrichia_montana_%28a,_164114-475524%29_9422") |
_5019 with arrow.JPG "Ptilium_crista-castrensis_%28c,_113329-471504%29_5019%20with%20arrow") |
| Hedwigia ciliata, leaf without a costa | Syntrichia montana, a single costa emerging as a long hyaline awn | Ptilium crista-castrensis, short double costa marked with red arrows |
According to their anatomy, homogenous and heterogenous costa are distinguished. In homogenous costa no internal cell differentiation is seen. This type is found in pleurocarpous Bryidae, Andreidae and some Grimmiales and Orthotrichales [4, p. 80]. Heterogenous costae are built from epidermal cells, stereids, deuter cells and/or hydroids and leptoids. In Polytrichidae deuter cells, hydroids and leptoids are flanked by stereids, in Dicranales, Encalyptales and Pottiales no leptoids are found. The costae of Leucobryaceae can cover half of the width of the leaf and is built from chloro- and hyalocysts [4, p. 82].
On the costae of the Polytrichaceae green lamellae are found, running from the apex down to the sheathing base of the leaf. The margins are covered with a hydrophobic wax to prevent flooding with water, which would reduce CO2 absorption. In Dicranaceae ridges are found on the dorsal portion of the costa. Another very special morphology is found in Pilotrichum. Its costa forms crests which can produce propagules [1, p. 68 et. seqq].
Pictures from Hermann Schachner, CC0, via WikimediaCommons. Anatomy interpreted with [4, p. 81]. Edited by Simon Hofer.
_9263.JPG "Schistidium_rivulare_%28a,_144638-481306%29_9263") |
_7931with markings.JPG "Mnium_lycopodioides_%28a,_144805-474432%29_7931with%20markings") |
_1215 with markings.JPG "Polytrichastrum_longisetum_%28b,_144747-474405%29_1215%20with%20markings") |
| Schistidum rivulare, homogenous costa. No differentiation recognizable | Mnium lycopodioides, heterogenous costa. the center of the costa is formed by sterides (St). Note the multistratose margin. | Polytrichastrum longisetum. A large and complicated heterogenous costa. Two bands of sterides (St) encase deuter cells (D), hydroids (H) and leptoids (L). Note the lamellae. |
Leucocysts
Leucocysts, also called Hyalocysts, are cells devoid of cytoplasmatic content. It is assumed their main purpose lies in water retention. Mosses mostly lack the mechanisms used by tracheophytes to control their water balance, in hot conditions they simply desiccate. Leucocysts as dedicated water storage could extend the time a moss can be photosynthetically active [1, S. 70]. In Sphagnum, leucocysts together with chlorophyllose cells form the lamina of a leaf. The pattern of their arrangement is diagnostically useful. Other properties of sphagnum leucocysts are wall thickenings and pores opening the lumen towards the environment. Their formation is suppressed when the moss is grown submerged in low light conditions [1, p. 71]. In Leucobryaceae layers of chlorophyllose cells are sandwiched between one or multiple layers of leucocysts. Pores connect adjacent leucocysts. Whether these also serve as water storage is unsure. The created water envelopewould greatly reduce CO2 diffusion to the chlorophyllose cells. Robinson (1985) assumes on juvenile leaves the leucocysts store air and thus CO2 for assimilation and on mature leves they store water [1, p. 71]. In some Pottiaceae and in Encalypta hyaline basal cells are found, for which a role in water storage and conduction is assumed [1, S. 71].
Pictures by Hermann Schachner, CC0, via WikimediaCommons
_4844.JPG "Sphagnum_cuspidatum_%28c,_150948-474700%29_4844") |
_7354.JPG "800px-Leucobryum_glaucum_%28c,_145632-482732%29_7354") |
_1582.JPG "Weissia_longifolia_%28a,_153208-482342%29_1582") |
| Sphagnum cuspidatum, lamina formed from a mosaic of hyaline leucocysts and chlorophyllose chlorocysts typical for the genus Sphagnum. | Leucobryum glaucum, leaf cross section - note the visible pores. | Weissia longifolia, base of a leaf, showing the transition of hyaline to chlorophyllose cells. |
Sex organs - Archegonia and Antheridia
Generally, mosses are capable of sexual reproduction. Female gametangia are called archegonia, male antheridia. Together with paraphyses, sterile unbranched filaments and special leaves the gametangia form budlike or rosulate structures, called perichaetium (male) or perigonium (female) [1, S. 74, 4, S. 88].
The sex of a plant is likely genetically determined. In some mosses chromosomes analogous to the XY-chromosomes of humans (heteromorphic and bivalent) can be found, which in some cases correlate with the sex of the plant. Expression, however, is more complicated: Not every plant expresses sex organs and expression varies on short distances and between the sexes of a species. How chromosomes determine the sex and which environmental factors lead to expression of gametangia is not understood [1, S. 74].
Mosses can be bisexual (monoicy), bearing both gametangia on a single plant, or unisexual (dioicy), where a plant only expresses either one. In monoicy, several arrangements are differentiated: Synoicy, antheridia and archegonia are mixed in one cluster, paroicy, archegonia surrounded by antheridia, and autoicy, where both gametangia are separated into their own clusters. A rare and interesting variant is pseudodioicy, in which the same plant expresses different gametangia at various times [1, S. 75].
Picture by Simon Hofer, CC SA-BY 4.0

Sexual dimorphism in dioicous species is rare. In phyllodioicous or pseudoautoicous taxa males are reduced to a single bud emerging directly from protonema growing as an epiphyte on the female plant [1, S. 75].
References
[1] Goffinet, B. u. Shaw, A. J.: Bryophyte biology. Cambridge, New York: Cambridge University Press 2009
[2] Vanderpoorten, A. u. Goffinet, B.: Introduction to bryophytes. Cambridge, UK, New York: Cambridge University Press 2009
[3] Cohem, K. M., Finney, S. C., Gibbard, P. L. u. Fan, J.-X.: The ICS International Chronostratigraphic Chart. Episodes 36: 199-204. 2013, updated
[4] Frahm, J.-P.: Biologie der Moose. Berlin, Heidelberg: Springer Berlin Heidelberg 2001
[5] Esser, K.: Kryptogamen II Moose · Farne. Praktikum und Lehrbuch. Berlin, Heidelberg: Springer Berlin Heidelberg 1992
[6] Frahm, J.-P., Frey, W. u. Döring, J.: Moosflora. UTB, Bd. 1250. Stuttgart: Ulmer 2004
[7] Malcolm, B. u. Malcolm, N.: Mosses and other bryophytes. An illustrated glossary. Nelson: Micro-Optics Pr 2000